11.1 Soil-Plant-Atmosphere Continuum
We need to understand transpiration and root water uptake within the context of the soil-plant-atmosphere continuum, the continuous pathway by which water moves from the soil, through plants, to the atmosphere. Water moves along this pathway in response to the typically large difference in water potential between the soil and the atmosphere. For example, in a typical greenhouse in Shaanxi, China, the air had a water potential of -168 MPa while the soil water potential was -0.26 Mpa (Fig. 11‑1, left side). When the air in the greenhouse was humidified using a fogging system, the water potential of the air rose to -82 MPa (Fig. 11‑1, right side), while the soil water potential was essentially unchanged. As a result the water potential difference driving transpiration was reduced by ~50% and the cumulative transpiration of the tomato plants being grown in the greenhouse was reduced by 20% [1]. This example illustrates that transpiration is, in one sense, not an active physiological function of plants but rather a primarily passive concession plants must make to the almost insatiable atmospheric demand for water.
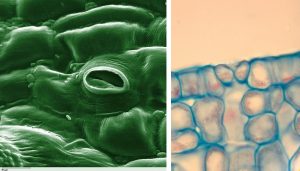
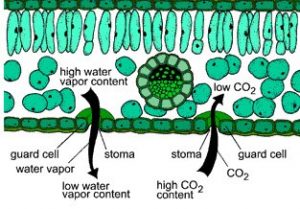
To understand the physics of transpiration, we need to consider the processes occurring at the stomatal level. Plant stomates are pores or openings on the exterior surfaces of leaves and other plant organs (Fig. 11‑2, left side). A pair of specialized cells, called guard cells, allow the plant to open and close the stomates (Fig. 11‑2, right side). The opening and closing of the stomates regulate the flow of gases such as water vapor and carbon dioxide into and out of the plant (Fig. 11‑3). The plant must open the stomates to take in carbon dioxide from the atmosphere, the first step in the process of photosynthesis. But in so doing, the plant unavoidably loses water vapor to the atmosphere. The oxygen gas generated in the process of photosynthesis also exits the plant through the stomates.